
L'arca olearia
L'origine dell'addomesticazione dell'olivo: tutto inizia nel Pliocene

Vi è stata una continua mescolanza genetica, con popolazioni selvatiche presenti nei territori dell'Europa meridionale e dell'Africa settentrionale, spesso coinvolgendo, in questo mixage, le popolazioni selvatiche geneticamente ricche
18 giugno 2021 | Alessandro Vujovic
C'è una grande quantità di prove che indicano il Mediterraneo (MD) orientale come la culla del primo evento di addomesticamento della pianta dell’olivo coltivato (Olea europaea subsp. europaea var. europaea, L) partendo dall’olivastro (Olea europaea subsp. europaea var. sylvestris, Mill. Lehr). Secondo studi archeologici, paleobotanici e genetici, l’olivo è stato addomesticato da progenitori selvatici circa 6.000 anni fa dai popoli camito-semitici.
Le analisi filogenomiche mostrano un processo secondario, continuo, di addomesticamento che coinvolge, oltre l’evento iniziale nel Levante, il prosieguo di eventi ricorrenti di mescolanza genetica indipendente, con popolazioni selvatiche presenti nei territori dell'Europa meridionale e dell'Africa settentrionale, spesso coinvolgendo, in questo mixage, le popolazioni selvatiche geneticamente ricche.
Irene Julca (2020) riferisce che l’olivo coltivato mostra livelli di diversità genetica leggermente inferiori rispetto alle forme selvatiche, il che può essere spiegato dal verificarsi di un collo di bottiglia della popolazione risalente a 3.000-14.000 anni fa durante il periodo di addomesticamento primario, seguito da una ricorrente “introgressione” da parte delle popolazioni selvatiche. Viene definito “collo di bottiglia” quando c’è una diminuzione delle frequenze alleliche (cioè una “minore diversità genetica”), in risposta ad eventi ambientali casuali, in modo tale che sopravvivono pochi individui selezionati della popolazione.
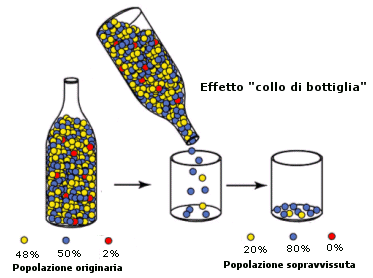
Storicamente fin dall'antichità, le cultivar sono state piantate vicino alle piante selvatiche, in questo modo si sono scambiati i pollini che hanno portato ad un'efficace produzione delle colture e ad una ibridazione storica.
In riferimento alla domesticazione dell’olivastro, nelle cultivar, i geni che sono associati alla risposta a stress ed a processi di sviluppo, sono stati selezionati positivamente, ma non ci sono prove che i geni coinvolti nella dimensione dei frutti / semi o del contenuto di olio fossero sotto selezione positiva, anche se gli studiosi prevedevano fossero stati influenzati dall'addomesticamento. Però, nelle forme addomesticate, potrebbe essere cambiata l’espressività epigenetica, cioè un apporto di cambiamenti fenotipici ereditabili senza variazioni del genotipo.
Gli studi di Besnard G. (2013), basati su marcatori plastidici e nucleari, hanno suggerito scenari di addomesticamento controversi ma non necessariamente incompatibili. a causa della bassa diversità dei plastidi. Ad esempio, la quasi totalità delle cultivar di olivo di tutto il MD condivide lo stesso aplotipo plastidico "orientale".
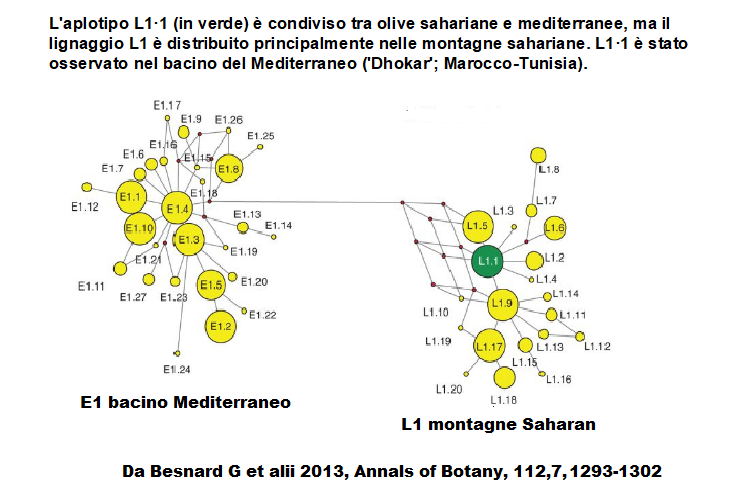
Sono riconosciute quattro linee di aplotipi plastidiali (ptDNA): tre dal bacino del MD (E1, E2, E3) e una dal Sahara (L1).
L'aplotipo L1·1 è condiviso tra olive sahariane e mediterranee; il lignaggio L1 è distribuito principalmente nelle montagne sahariane e un solo aplotipo è condiviso tra la sottospecie sahariana laperrinei e la europaea, ed è stato rilevato in una cultivar del Maghreb, la cv "Dhokar".
I microsatelliti nucleari mostrano modelli geografici di differenziazione genetica nell'olivo MD che riflettono le origini primarie delle cultivar nel Levante e indicano un'elevata differenziazione genetica tra europaea e laperrinei. Non è stato rilevato alcun ibrido di prima generazione tra europaea e laperrinei, ma in alcune adesioni, tra cui "Dhokar", è stata riscontrata una commistione reciproca “recente” tra sottospecie mediterranee e sahariane.
Secondo alcuni autori il lignaggio della subsp. Sylvestris mostra, dal ptDNA, una differenziazione in tre linee aplotipiche originate nel periodo compreso tra il Pliocene (5,32 Ma -1,78 Ma BP; Ma= milioni di anni, BP=Before Present) e il Pleistocene (2,58 Ma BP-11.700 BP) prima dell'ultima Grande Glaciazione, pertanto i bacini genetici sono due: uno relazionabile all’areale orientale (E1), limitato ad est della Grecia, mentre il lignaggio plastidico (E2, E3) è legato alle regioni centrali ed occidentali del MD.
Secondo Besnard G. (2013b) dei tre clorotipi E1/E2/E3, quello comune agli olivi selvatici che appartiene alla linea orientale E1, caratterizza il 90% delle cultivar ed è presente nei cloroplasti degli olivi selvatici di tutto il bacino del MD.
La relazione tra questi gruppi ha mostrato anche due caratteristiche interessanti: il gruppo occidentale E3 (Spagna meridionale e Portogallo) conserva l'impronta digitale di un collo di bottiglia genetico e, sorprendentemente, è strettamente correlato alle forme coltivate nel Levante.
Diez C.M. (2015) ha ipotizzato che nelle cultivar del MD centrale, il 20% della diversità genetiche potrebbe essere attribuita ad una acquisizione tramite introgressione con materiale genetico delle popolazioni selvatiche locali.
Dalle ricerche del gruppo di Irene Julca (2018, 2020), si evidenzia che la storia evolutiva dell'olivo comprende non solo una duplicazione dell’intero genoma, specie-specifico, ma anche due eventi di allopoliploidizzazione ancestrali (paleoploidizzazione), cioè un raddoppio del corredo cromosomico tale da aver contribuito ad una maggiore resistenza a condizioni avverse ed alla domesticazione.
Dopo la poliploidizzazione, il genoma può tornare progressivamente a uno stato diploide. Il genoma della varietà sylvestris e della varietà europaea ha una dimensione diploide distribuita in 46 cromosomi (2n), ma attualmente si conoscono, come poliploidi, l’Olea europaea subsp. cerasiformis (4n) e la subsp. maroccana (6n).
Tra oltre 1000 geni che sono assenti nel genoma sylvestris, ma presenti nella forma coltivata, ci sono funzioni associate alla risposta allo stress, come: HIPP (proteine ​​vegetali isoprenilate associate a metalli pesanti), LEA (embriogenesi tardiva abbondante), legante all’acido salicilico o altri geni associati alla crescita e allo sviluppo, come il RALF (fattore di alcalinizzazione rapida), oppure la caffeoil-scichimicato esterasi (CSE), che è un enzima nella via biosintetica della lignina.
I genomi dei plastidi sono congruenti con tre principali lignaggi (E1, E2 ed E3) già descritti e, poiché gli organelli nell'olivo sono ereditati dalla madre, i risultati suggeriscono che le cultivar e gli individui selvatici condividono un antenato materno comune.
Inoltre, sia Julca I. (2020) che Jiménez-Ruiz J. (2020) hanno trovato una relazione molto stretta a livello organellare e nucleare tra le cultivar orientali della Siria, Iran, Libano e Turchia e quelle della Spagna-Portogallo, con un collo di bottiglia locale probabilmente correlato all'introduzione del germoplasma dell'olivo nella Spagna meridionale durante il periodo musulmano. Questo periodo, iniziato nel 700 d.C., durò otto secoli e forse rimodellò il germoplasma dell'olivo coltivato della penisola iberica a causa della migrazione di cultivar dal Levante con la sostituzione del germoplasma locale precedentemente coltivato.
I risultati di varie ricerche suggeriscono che la linea materna di cultivar con aplotipo plastidico E3 provenisse da olive selvatiche del MD occidentale, mentre la linea paterna proveniva principalmente da individui addomesticati del bacino del MD orientale.
È importante sottolineare che il livello di introgressione è ampiamente indipendente dall'aplotipo plastidico (tutte le cultivar degli aplotipi plastidi E2 ed E3 e alcuni di E1 ne mostrano segni), suggerendo che, in alcuni casi, si sono verificate diverse introgressioni nei lignaggi della madre e del padre, indipendentemente.
L'origine del materiale genetico introgresso può essere meglio dedotta quando il lignaggio del donatore può essere ricondotto attraverso la linea materna. Ciò è particolarmente vero per gli individui campionati da regioni vicine all'origine dell'addomesticamento, come le cultivar dalla Siria, Iran, Turchia e Libano e gli olivastri dalla Turchia, che mostrano pochi o nessun segno di introgressione.
I geni selezionati positivamente nelle olive coltivate sono associati alla risposta a stress biotici e abiotici ma non ci sono prove di selezione di geni associati alla dimensione dei frutti o al contenuto di olio.
Potrebbero interessarti
L'arca olearia
I soliti ignoti dell’oliveto: quando il nemico non è la mosca dell'olivo

Cecidomia, tignola carpofaga, margaronia e altri fitofagi minori stanno emergendo come protagonisti di danni ingenti, spesso sottovalutati. L’olivicoltore deve aggiornare il proprio “libretto” dei nemici per proteggere la produzione da minacce silenziose ma sempre più insidiose
27 luglio 2026 | 14:00
L'arca olearia
Enzimi e temperatura di gramolazione: nuove strategie per valorizzare colore e qualità dell’olio extravergine di oliva

Uno studio condotto su tre varietà italiane evidenzia come gli enzimi Bioliva e Rapidase Adex D, insieme alla gestione della temperatura di gramolazione, possano influenzare il profilo cromatico, la presenza di sostanze funzionali e alcuni parametri legati alla stabilità e alla qualità sensoriale dell’olio
25 luglio 2026 | 12:00
L'arca olearia
Congelamento convenzionale e criogenico delle olive: effetti sulla qualità dell'olio varietale

Il congelamento convenzionale è economicamente sostenibile, con un incremento del prezzo finale dell'olio dello 0,6% per 7 giorni e del 5% per un anno, mentre il criocongelamento renderebbe il prodotto circa 4-5 volte più costoso, pur preservando meglio i composti fenolici e l'attività antiossidante
24 luglio 2026 | 15:00
L'arca olearia
Le differenze nell'olio di oliva prodotto da olivi giovani e secolari svelati dalla risonanza magnetica

Un recente studio basato sulla spettroscopia NMR ha evidenziato differenze significative nella composizione chimica dell'olio di Peranzana in funzione dell'età degli alberi, offrendo nuovi spunti per la valorizzazione degli olivicoli secolari pugliesi
24 luglio 2026 | 14:00
L'arca olearia
La cecidomia dell'olivo e le sue alleate: comprendere il tritrofismo per una difesa sostenibile

La Dasineura oleae è un insetto galligeno che negli ultimi anni sta causando preoccupanti infestazioni in tutto il Mediterraneo. L'attacco della cecidomia modifica profondamente la fisiologia e l'anatomia delle foglie, riducendo la fotosintesi fino al 35% e la conduttanza stomatica del 28%
24 luglio 2026 | 13:00
L'arca olearia
La rivoluzione silenziosa nell'estrazione dell'olio di oliva: il progetto IT4NUEVOO

Il progetto IT4NUEVOO ha sviluppato tecnologie innovative per l'estrazione di un olio con un contenuto ancora più elevato di sostanze salutari, ponendo particolare attenzione alla preservazione dei composti fenolici durante il processo produttivo
24 luglio 2026 | 12:00




Commenta la notizia
Per commentare gli articoli è necessario essere registrati
Accedi o Registrati